Autores
Luiz Fernando Jobim (1), Maria Regina Jobim (2), Gisele Ewald (2), Mariana Jobim Wilson (3), Beatriz Chamum (4).
1. Doutor em Medicina pela UFRGS, Professor da Faculdade de Medicina da UFRGS e chefe do Serviço de Imunologia do Hospital de Clínicas de Porto Alegre. Pós-graduação na Universidade do Texas e de Oxford.
2. Mestre em Genética e Biologia Molecular pela UFRGS.
3. Mestre em Medicina pela UFRGS e médica do Serviço de Imunologia do Hospital de Clínicas de Porto Alegre.
4. Bióloga do Serviço de Imunologia do Hospital de Clínicas de Porto Alegre
Resumo
Novos testes de DNA foram desenvolvidos após a queda das torres gêmeas de Nova York, permitindo a identificação de DNA degradado, fato que acontece após o falecimento devido à decomposição dos corpos pela umidade, tempo, excesso de calor, entre outros fatores. Nesse artigo, os autores apresentam os conhecimentos atuais e as precauções necessárias para perícias onde o suposto pai é falecido, como a identificação de quem deve ser intimado para o exame de DNA, de acordo com o grau de importância na reconstituição do falecido, levando em conta as pessoas disponíveis para análise pericial. A identificação humana pelo estudo do DNA do cromossomo masculino Y e X é abordada como parte dos novos testes de DNA disponíveis para a investigação de paternidade, especialmente quando o suposto pai é falecido. Um caso de perfeita identificação do perfil genético do réu, após 28 anos de seu falecimento é analisado, permitindo o conhecimento de como é possível obter-se resultados definitivos à luz dos novos testes de mini-STRs do DNA.
Introdução
A identificação de paternidade, nos casos onde o suposto pai é falecido, pode sofrer limitações pela escassez de indivíduos para análise, tanto na parte autora, quanto na do suposto pai. As razões podem ser várias, como o falecimento de uma das partes, a falta de localização de outra, a ausência do país ou por não desejarem realizar o exame pericial. Na maioria desses casos, acontecem questionamentos sobre “quem deverá ser intimado” para a realização dos testes de reconstrução do DNA do suposto pai (JOBIM, 1995).
O DNA humano está disponível em quantidade suficiente para análise em todas as células nucleadas do sangue e tecidos, em menor concentração no soro ou no plasma e no líquido amniótico placentário (urina da criança). No cabelo, ele está bem representado nos bulbos capilares, enquanto que nos ossos fica enclausurado nas células conhecidas como osteoclastos. Em indivíduos falecidos há poucas horas ou dias, podemos recolher facilmente amostras de sangue puncionando veias ou o próprio coração, além de cabelos que devem conter a raiz (um número não inferior a vinte fios). Com o passar do tempo, até quatro semanas da morte, podemos usar amostras de músculo esquelético (um centímetro quadrado). Quando o indivíduo faleceu há mais tempo, os cabelos já perderam a raiz e os tecidos remanescentes costumam ter o DNA degradado, não sendo adequados. Nesses casos, os ossos e os dentes são de grande importância. Os últimos podem ser decisivos se houve morte por fogo, pois a raiz dentária fica protegida, até certo ponto, pela estrutura dentária. O fêmur e os dentes molares e pré-molares costumam ser as peças preferidas após a exumação de indivíduos falecidos para a finalidade de análise do DNA (JOBIM, 1998). É possível também identificar pessoas falecidas pela análise de antigas escovas de dente, a partir da extração do DNA de células bucais que ficam com seu esqueleto preso nas cerdas, possibilitando testes confiáveis (JOBIM, 2004).
O DNA humano pode ter origem autossômica, ou seja, proveniente de cromossomos não sexuais, assim como pode existir DNA do cromossomo masculino Y ou do feminino X. A maioria dos testes atuais analisa diversos locos ou locais cromossômicos de DNA autossômicos, entretanto os testes de DNA do cromossomo Y e X começam a ter sua importância em determinados tipos de perícias.
Os primeiros genes marcadores do DNA humano foram descritos por Jeffreys e colaboradores, sendo conhecidos como minissatélites. Esses são locos ou regiões onde acontecem repetições de conjuntos de oligonucleotídios em “tandem”, ou seja, consecutivos ou lado a lado. Essas repetições têm importância médica somente por serem marcadores biológicos da espécie humana. Os minissatélites não são mais utilizados atualmente devido a sua substituição pelos microssatélites ou STRs (“short tandem repeats” ou repetições curtas em tandem”). Esses locos e seus genes são identificados pelo método de polimerase chain reaction (PCR), (MULLIS, 1987).
Analisando-se um número suficiente de locos genéticos de STRs podemos concluir pela inclusão da paternidade ou pela sua exclusão. A última acontece quando um marcador (alelo) do DNA do autor não existir no perfil do DNA paterno. No caso da existência dessa inconformidade podemos estar frente a uma exclusão de paternidade ou pode ter acontecido uma mutação. Os laboratórios, na maioria das vezes, concluem pela exclusão somente após a identificação de mais de dois locos onde o alelo paterno do filho não é compartilhado pelo suposto pai.
Quando o suposto pai é falecido, procuramos reconstituir ou identificar os alelos paternos pela análise de ascendentes ou descendentes do mesmo. A mãe do autor e a viúva do suposto pai são sempre importantes e devem ser incluídas, sendo que a última deve ser intimada quando os seus filhos com o falecido (supostos meio irmãos) são analisados. A razão é que identificamos no autor (em cada loco do DNA) o alelo materno e o segundo alelo é obrigatoriamente paterno, servindo para comparação com os alelos paternos identificados nos parentes do falecido. Quando analisamos os supostos avós paternos, não necessitamos da viúva, pois o DNA do falecido estará sempre presente em um dos pais. A ausência de um alelo em comum entre o autor e seus supostos avós é indicativa de exclusão de paternidade, embora outras exclusões devam ser identificadas para a certeza científica.
Mesmo em casos onde as mães não realizam os testes, pretendemos identificar um alelo do autor em comum com um de seus possíveis tios ou irmãos paternos. Nesses casos, a certeza diminui, pois o autor tem dois alelos por loco e estamos considerando os dois alelos ao mesmo tempo, pois não sabemos qual é o alelo paterno. Nos cálculos matemáticos, esse fato induz a resultados com índice e probabilidade de paternidade menores. Uma das regras básicas para o entendimento dos casos periciais onde o suposto pai é falecido é que cada indivíduo apresenta no máximo dois alelos por loco de DNA (heterozigoto). Caso apresentar somente um é porque herdou o mesmo alelo de seu pai e de sua mãe (homozigoto). Tendo isso em mente, entre supostos irmãos (quando não analisamos a respectivas mães) só é possível existir quatro alelos diferentes, dois de herança materna e dois paterna. Um quinto alelo diferente, no autor, representa a exclusão da paternidade. Caso analisarmos a viúva e seus filhos com o falecido, poderemos identificar ou reconstruir o perfil dos dois alelos do mesmo. Nessa situação, um terceiro alelo no autor exclui a paternidade.
Uma das dificuldades encontradas em testes de paternidade é a possibilidade de que um irmão do suposto pai seja o verdadeiro pai biológico do autor. Nesse caso é possível calcularmos matematicamente essa hipótese. A fórmula fornece o resultado do índice avuncular e permite o cálculo da probabilidade cumulativa avuncular, ou seja, a probabilidade do homem testado ser tio do autor. Podemos também conhecer quantas vezes o homem testado tem de chances de ser o verdadeiro pai do que ser tio do autor.
Preferências na Análise do DNA com suposto pai falecido
a) O autor, sua mãe (se existente) e ambos os supostos avós paternos.
b) O autor, sua mãe, a viúva (se existente) e seus filhos com o falecido, além de um dos avós paternos (se existente). Quanto maior o número de filhos melhor a possibilidade de resultados conclusivos. Em casos onde existem muitos filhos, iniciamos com a análise de três ou quatro. Na eventualidade de somente existir um, incluir um dos avós e supostos tios (irmãos do falecido).
c) O autor, sua mãe, suposta avó e filhos (irmãos do suposto pai – maior número possível).
d) O DNA do cromossomo Y deverá ser analisado em casos onde não se conseguiu uma exclusão definitiva, existindo mutações ou a probabilidade positiva de paternidade não alcançou pelo menos 99 %. Dessa maneira podemos identificar se o autor e o suposto pai pertencem à mesma família de homens. Além do autor, podemos analisar comparativamente outros indivíduos masculinos pertencentes à mesma linhagem do falecido suposto pai. Entre eles, um filho, um irmão, um primo (filho de um irmão do falecido), um neto (filho de um filho do falecido) entre outros. O importante é que a corrente de indivíduos seja sempre de homens, não podendo ser intercalada por uma mulher.
e) A análise do DNA do cromossomo X poderá ser utilizada em casos difíceis onde não se conseguem exclusões e os índices e probabilidades são baixos (menos de 99%). Nem todos os casos podem ser avaliados pelo DNA do X. Nos casos onde existe somente a autora e uma suposta irmã, existirá sempre um alelo em comum entre as duas se forem filhas do mesmo pai. Isso se deve ao fato que o pai biológico sempre passa o mesmo DNA do X para suas filhas, o que não acontece para os filhos homens, quando passa o próprio DNA do Y. É crescente a utilização desses marcadores como veremos mais adiante.
f) Outra opção seria a da análise do autor e de um suposto irmão. Nesse caso nunca será obtida a exclusão da paternidade e a parte ré estará sendo prejudicada. Isso é devido ao fato de que existindo dois alelos por loco, o autor poderá ter dois alelos detectados. O suposto irmão poderá ter um dos alelos com identidade com o autor, mas esse pode ser de origem materna e não de seu pai biológico. Além disso, quando não existirem identidades, podemos estar frente a dois alelos diferentes do suposto pai presentes um em cada filho, o que não exclui a paternidade, mas também não inclui. O mais lógico, em nossa opinião é observarmos o valor dos índices e adicionarmos a análise do DNA do Y se forem dois supostos irmãos. Nesse caso, podemos excluir a paternidade ou afirmar que somente pertencem a mesma família de homens somente.
g) A exumação do cadáver do suposto pai é necessária sempre que não existam ascendentes ou descendentes ou quando esses são escassos, ou os resultados obtidos com os métodos anteriores (indiretos) não alcançou probabilidades de paternidade aceitáveis ao perito e ao juiz. Nesses casos, podemos analisar o DNA do autor e o extraído diretamente do suposto pai. No caso de dúvida que o cadáver pertença ao suposto pai, podemos incluir um reconhecido filho biológico na perícia. Também devemos ter cautela, pois já participamos de casos onde incluíram uma filha registrada que a família sabia não ser filha do suposto pai.
h) A exumação deverá ser realizada por médico legista ou pelo perito. As partes devem comparecer no local da exumação. Os novos métodos que utilizam os mini-STRs devem ser utilizados. A extração do DNA deve ter qualidade, pois caso contrário, os resultados não serão confiáveis. As imagens ou eletroesferogramas dos perfis genéticos dos indivíduos devem ser incluídas no laudo, pois permitem análise pelo juiz e pelos assistentes técnicos (Fig. 2).
Avaliação do DNA pela reação em cadeia da polimerase (PCR)
A PCR é um método extremamente poderoso na avaliação da individualidade humana. Ela permite a amplificação seletiva, in vitro, de seqüências específicas do DNA alvo que se deseja estudar. O princípio da reação está no conhecimento da seqüência do DNA alvo e em reação bioquímica complexa. A ciência básica forneceu as seqüências das regiões de milhares de microssatélites de importância em genética forense. Os locos de microssatélites conhecidos como STRs apresentam alelos ou fragmentos de DNA de 100 a 400 pares de bases. Esses são os preferidos na análise pericial de casos de identificação humana. Os STRs permitem a amplificação de milhares de cópias de fragmentos específicos de DNA, sendo o método utilizado na maioria dos laboratórios (GONZÁLEZ-ANDRADE, 2008).
A identificação dos fragmentos amplificados é realizada após eletroforese capilar, seguida pela detecção de sinal fluorescente. O sinal fluorescente acontece quando marcamos os primers com fluorocromos. Dessa maneira podemos usar analisadores de DNA (seqüenciadores automáticos) que identificam os fragmentos por intermédio de um laser. Um computador com programa dedicado permite observarmos curvas ou picos de DNA que são comparados entre as partes (Fig. 1, 2 e 3).

Após a amplificação de DNA utilizamos uma escada alélica (ladder) na identificação de fragmentos na reação de PCR. Todas as possibilidades de alelos dos locos amplificados podem ser observadas e comparadas com o produto amplificado. Dessa maneira, é possível conhecer o tamanho do fragmento em pares de base (pb) e seu correspondente alelo existente no indivíduo analisado. Os casos de exumação devem ser analisados com reagentes conhecidos como mini-STRs.
Os STRs são muito utilizados na análise de investigação de paternidade, devendo-se analisar um número suficiente de locos, geralmente 12 ou mais locos. Os STRs e mini-STRs são valiosos no estudo de casos em que exista necessidade de análise de ossos, dentes, fios de cabelo, manchas de sangue, entre outros (MAYNTZ-PRESS, 2004).
A técnica da PCR permite também a identificação do sexo do indivíduo. O loco da amelogenina apresenta um alelo de 106 pares de bases nas mulheres (XX) e um de 106 e outro de 112 pares de bases nos homens (XY).

Figura 2. Caso simples de avaliação da paternidade com amostras da autora, viúva e dois supostos meio irmãos da autora. No loco D21S11, a possível irmã da autora (PI1) herdou o alelo 31,2 de sua mãe (viúva), sendo que o alelo 31 é de origem paterna e ausente no autor. Completa esse resultado a análise do possível irmão da autora (abaixo) que herdou o alelo 29 de sua mãe (viúva), sendo que o alelo 28 é de origem paterna e ausente no autor. Nesse caso, identificamos os dois alelos paternos do falecido (31 e 28), excluindo a paternidade nesse loco.

Figura 3. Caso de exclusão de maternidade pela análise do DNA extraído da escova de dente da falecida mãe e do DNA de seu suposto filho. Observam-se exclusões nos locos D16S539 onde a mãe apresenta os alelos 9 e 11 e o pretenso filho, os alelos 12 e 14. No loco D2S1338 a mãe apresenta alelos 17 e 22 e o suposto filho os alelos 19 e 24.
STRs do Cromossomo Y
Recentemente foram descritos STRs no cromossomo masculino Y. Algumas características desse cromossomo fazem com que seus microssatélites (STRs) sejam importantes na análise forense do DNA (PONTES, 2006). Devido à falta de um cromossomo homólogo, não existe recombinação durante a meiose. Sendo assim, só identificamos alelos de origem masculina, herdados em bloco dos antepassados masculinos. A herança em bloco de alelos de diferentes STRs do mesmo cromossomo é denominada herança haplotípica e o conjunto dos alelos chama-se haplótipos (BUTLER, 2002 e 2003).
Dessa maneira, é possível a identificação de diversos locos de STRs em um homem, sendo os mesmos alelos encontrados em seu pai, irmãos, tios, primos, avô e demais antepassados masculinos. Estudos apontam um baixo índice de mutação, podendo os mesmos haplótipos ser encontrados em várias gerações de homens, alcançando talvez várias centenas de anos (GILL, 2001).
Cada alelo ou marcador do Y de determinado local do DNA apresenta uma freqüência na população, existindo bancos de dados especializados (www.ystr.org/index.html). Se examinarmos sete ou mais locos do DNA do Y, poderemos conhecer matematicamente a existência ou não de uma relação genética entre indivíduos, no entanto nunca poderemos afirmar o grau de parentesco (KAISER, 1994).
Os STRs do cromossomo Y permitem reconhecer a exclusão de paternidade quando o possível pai é falecido, analisando-se o pretenso filho e homens aparentados com o suposto pai (Fig. 4).
Os principais locos de STR do Y são: DYS19, DYS389 I, DYS389 II, DYS390, DYS391, DYS392, DYS393, GATA H4, DYS437, DYS438 e DYS448 (MAYNTZ-PRESS, 2007).

Figura 4. Exclusão de paternidade pelo DNA do cromossomo Y entre o autor (João Fictício) e um neto de seu suposto pai (José Fictício). No loco DYS437 o autor apresenta o alelo 16 e o José Fictício o alelo 14. No loco DYS438, um apresenta o alelo 9 e o outro o alelo 12. No loco DYS448 as diferenças são devidas aos alelos 19 e 18.
Um estudo realizado pelo Prof. Chris Tyler-Smith, da Universidade de Oxford, permitiu a provável identificação do conjunto de genes do cromossomo Y de Gengis Khan. As amostras utilizadas foram da população masculina da Mongólia, levando-se em conta que o guerreiro mongol teve mais de cem filhos, os quais, por sua vez, tiveram outros milhares de filhos, todos com o mesmo perfil genético do cromossomo Y. Esse perfil é encontrado, na atualidade, em grande número de indivíduos mongóis do sexo masculino.
Outro estudo recente de importância foi a identificação de Sadaam Hussein, realizada pela comparação do conjunto ou haplótipo do DNA do cromossomo Y entre ele e o DNA extraído dos corpos de seus filhos mortos em combate (Fig. 5).

Figura 5. Sadaan e seus filhos analisados pelo DNA do Y – Buttler, J.M. (2005) em Forensic DNA Typing, 2a edição.
STRs do Cromossomo X
Como foi dito anteriormente, o cromossomo X apresenta também locos de microssatélites e que permitem conhecer a herança sob outra perspectiva. A mãe apresentará em cada loco dois alelos do DNA do X (XX), enquanto os homens sempre apresentarão um alelo desse DNA, pois o outro pertence ao DNA do Y (XY).
Como exemplo, podemos observar na tabela 1 os resultados do DNA do cromossomo X de uma perícia onde a análise de outros 18 locos de DNA autossômico chegaram a uma probabilidade de paternidade de 90%, o que não é satisfatório, pois existe 10% de chance de não paternidade. Nesse caso, o suposto pai era falecido e estudamos a autora, sua mãe e duas irmãs do falecido ou supostas tias da autora. Devemos levar em conta que entre irmãs somente podemos ter três alelos do cromossomo X, um de origem paterna e dois de origem materna.
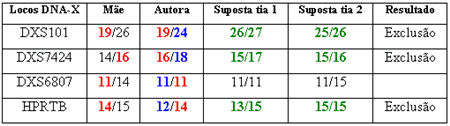
Tabela 1: Exclusão de paternidade pelo DNA do X de caso onde o suposto pai é falecido.
Nesse caso foi possível a identificação de três exclusões de paternidade. No loco DXS101, a autora herdou o alelo 19 de sua mãe, sendo o alelo 24 obrigatoriamente de origem paterna e que deveria estar presente no falecido réu, irmão das supostas tias. As tias apresentam o alelo 26 em comum, sendo esse alelo proveniente do pai ou suposto avô da autora (que só pode ter um alelo do cromossomo X). Logo, os alelos maternos presente nas tias são o 27 e 25. Os três alelos do cromossomo X possíveis de existir na família são os 26, 27 e 25. No entanto, a autora herdou o alelo paterno 24, o que é impossível de acontecer ou uma mutação ocorreu (Fig. 7).
No loco DXS7424, a autora herdou o alelo 16 de sua mãe e o 18 de seu pai biológico. As supostas tias herdaram o alelo 15 do seu pai e suposto avô da autora. Além disso, elas herdaram os alelos maternos 17 e 16. Os três alelos do cromossomo X possíveis de existir na família são 15, 17 e 16. No entanto, a autora herdou o alelo paterno 18, o que é impossível de acontecer ou outra mutação ocorreu. No loco HPRTB novamente aconteceu a exclusão da paternidade pelo DNA do X. O alelo paterno da autor é o 12 e as tias apresentam o alelo paterno 15 e dois alelos maternos 13 e 15.
Mini-STRs na Identificaçao de Material Degradado
Amostras de DNA degradadas sempre foram empecilhos na identificação de corpos após desastres ou longo tempo de falecimento. Os testes de identificação humana obtiveram o primeiro avanço na solução desses casos quando foi desenvolvida a reação em cadeia da polimerase (PCR). Esse já foi um avanço considerável, tendo em vista que os primeiros testes de DNA não amplificavam o mesmo, mas somente o identificavam.
No entanto, mesmo amplificando o DNA não se obtinham resultados adequados em amostras muito degradadas, pois nelas o DNA estava muito fragmentado devido a inúmeros fatores.
Na amplificação do DNA usamos reagentes que delimitam o local do DNA que queremos amplificar e, na maioria das vezes, os iniciadores ou “primers” amplificam regiões extensas do DNA. A novidade que permitiu os novos testes foi a redução do tamanho das regiões amplificadas, ou seja, reduzindo-se os fragmentos a serem detectados (COBLE, 2005 e 2006). Logo, com os novos reagentes podemos identificar os mesmos alelos do DNA, mas só que com tamanhos menores, permitindo definir alelos fragmentados ou degradados (DIXON 2006). Esses reagentes foram chamados de mini-STRs.
Como exemplo, um alelo de DNA que identificávamos pelo tamanho de 340 pares de bases, com os novos reagentes, podemos identificá-lo em 120 pares de base (DRABECK, 2004). Portanto é fácil o entendermos que estamos analisando o mesmo DNA, mas com o núcleo de identidade menor, o que chamamos de região de repetição (HILL, 2008). Sendo assim, a amostra identificada é de tamanho menor, sendo mais fácil ser detectada e comparada com a do autor (CHUNG, 2004).
Os primeiros mini-STRs foram desenvolvidos pelo Laboratório Bode (EUA) para a identificação das vítimas do World Trade Center, tendo suas seqüências de DNA publicadas em 2004. Os testes de mini-STRs são analisados em forma de “multiplex”, ou seja, é possível a amplificação e a análise de diversos locos de DNA ao mesmo tempo, devido à marcação dos iniciadores da reação (primers) com cores fluorescentes diversas (fluorocromos). Os testes são semelhantes aos que usamos para identificar outros STRs (BUTLER, 2003 e 2008).
Realizamos recentemente testes após a exumação do suposto pai, falecido há 28 anos. Utilizamos os mini-STRs (tabela 2, figuras 6 e 8). Os testes alcançaram um índice cumulativo de positivo de paternidade de 4874 e uma probabilidade cumulativa positiva de paternidade de 99,97%, sendo que em todos os locos analisados observamos alelos em comum entre a filha e o suposto pai (exumado).

Tabela 2. Resultados dos testes de DNA após a exumação do suposto pai. Em todos os locos observamos identidade entre alelos da autora e do suposto pai (Fig.8).
A complexidade do estudo de ossos reside principalmente na extração, purificação e uso de reagentes apropriados (Fig. 6). Os resultados atuais são nitidamente superiores aos que se obtiveram na década passada, obtendo-se um perfil genético no material exumado em 90% dos casos (Fig. 8). A contaminação existente nos ossos que se recebe para análise é danosa, tendo em vista à presença de agentes que degradam o DNA, assim como inibem a reação de amplificação. A terra, os fungos e as bactérias são os principais elementos indesejáveis e que podem acarretar diversos graus de degradação.

Os testes iniciam pela cuidadosa lavagem dos ossos para remoção de terra e outros materiais aderidos, seguindo-se da raspagem, com lixa, da superfície do osso até que fique claro ou limpo. Em seguida, usamos uma serra elétrica para separar fragmentos de cerca de 1 cm2. Esses são colocados em cilindros contendo fragmentos de metal e introduzidos num moinho eletromagnético com nitrogênio líquido. O instrumento é ligado e o magnetismo aciona a vibração do metal em alta velocidade, pulverizando os ossos. No final, teremos um pó de osso de cor branca. Quanto mais limpo o osso, melhor será a amplificação do DNA extraído de dentro dos osteoclastos. A razão da pulverização com nitrogênio líquido decorre da necessidade de baixas temperaturas para o DNA não ser destruído pelo calor.
O DNA deverá ser purificado antes de sua amplificação. Na maioria dos laboratórios utiliza-se o método de fenol/clorofórmio e precipitação do DNA da solução por álcool com posterior concentração em membrana ou filtro.
Considerações finais
A análise da individualidade humana iniciou com a tipagem de grupos sangüíneos, tipagem HLA (antígenos leucocitários humanos) e identificação dos minissatélites do DNA. Passou, então para os microssatélites ou STRs e chegou aos mini-STRs. Esses últimos são os ideais para a análise de ossos e material degradado, onde somente fragmentos pequenos de DNA são existentes, ao contrário do DNA original onde as moléculas encontram-se íntegras.
As tecnologias para análise do DNA estão em evolução. O próximo passo será a análise de pedaços de DNA conhecidos como SNPs ou polimorfismo de nucleotídeos de base única” (single nucleotide polimorphisms). Esses estão surgindo como marcadores de interesse para a comunidade forense devido: à sua abundância no genoma humano; ao seu baixo índice de mutação; por permitirem a análise de pequenos fragmentos de DNA e pela possibilidade de automatização das análises com tecnologias superiores (PHILLIPS, 2006).
A vantagem da utilização dos SNPs em relação aos STRs, se deve ao fato que os produtos de amplificação gerados são muito curtos (50 pares de base ou menos), permitindo o estudo de material degradado (como no caso de cadáveres em avançado de decomposição ou carbonizados). A tipagem de SNPs é complexa, existindo diversos métodos para a análise com fluorescência e luminescência. Um total de 50 locos de SNPs devem ser analisados para obter-se a equivalência ao estudo de 15 locos de microssatélites ou STRs. No entanto, em casos de extrema degradação poderemos aceitar um número menor de marcadores, tendo em vista ser a última possibilidade de análise genética (SANCHEZ, 2006 e VALLONE, 2006).

Figura 7. Estudo do DNA do cromossomo X (loco DX101). Caso explicado na tabela 1.

Figura 8. Eletroesferograma de parte do DNA (mini-STRs) do osso da fig. 6 e tab. 2.
